6.3.4. Marine Fish
Climatic factors affect the biotic and abiotic elements that influence the
numbers and distribution of fish species. Among the abiotic factors are water
temperature, salinity, nutrients, sea level, current conditions, and amount
of sea ice—all of which are likely to be affected by climate change. Biotic
factors include food availability and the presence and species composition of
competitors and predators. Clearly the relationship between climatic factors
and the fish-carrying capacity of the marine environment is complicated, although
water temperature can be used as a basis for forecasting the abundance and distribution
of many species (Lehodey et al., 1997). Water temperature also can have
a direct effect on spawning and survival of larvae and juveniles as well as
on fish growth, by acting on physiological processes. Sea temperature also affects
the biological production rate thus food availability in the ocean, which is
a powerful regulator of fish abundance and distribution.
The question of large-scale, long-term fluctuations in the abundance of marine
organisms, primarily those of considerable commercial importance, recently has
gained attention. Research has shown that variations (with cycles of 10-60
years or more) in the biomass volume of marine organisms depend on sea temperature
and climate (Ware, 1995). Examples include periodic fluctuations in the climate
and hydrographic regime of the Barents Sea, which have been reflected in variations
in commercial production over the past 100 years. Similarly, in the northwest
Atlantic Ocean results of fishing for cod during a period of 300 years (1600-1900)
showed a clear correlation between water temperature and catch, which also involved
changes in the population structure of cod over cycles of 50-60 years.
Shorter term variations in North Sea cod have been related to a combination
of overfishing and warming over the past 10 years (O'Brien et al.,
2000).
From 1987 to 1996, the world catch of all marine fishes averaged 74.5 Mt. From
1987 to 1993, catches were relatively stable, ranging between 71.6 and 75.9
Mt. There was a small increase over the period 1994-1996, ranging from
77.1 to 78.6 Mt (FAO, 1998). The 10 species with the largest landings represented
37.4% of the catch in 1996 and the next 10 species an additional 10.9%. Fluctuations
in abundance of species representing the 10 largest landings often have been
considered to result from overfishing and occasionally from a combination of
ocean environment changes and fishing effects. However, there is increasing
evidence to suggest that the impacts of climate variations are also having an
important effect (O'Brien et al., 2000).
|
Box 6-1. Regimes and Regime Shifts: Salmon and Sardine
Catch
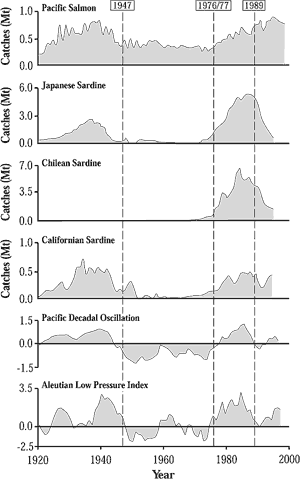
As the figure to the left shows, catches (Mt) of Pacific salmon (Oncorhynchus
sp.) and sardines (Sardinops sp.) fluctuate synchronously with large-scale
climate-ocean changes in the North Pacific, as indicated by the Aleutian
Low Pressure Index and SST as expressed in the Pacific Decadal Oscillation
both of which are shown as a 5-year running average (data from Beamish
et al., 1999, and Mantua et al., 1997). Persistent states or regimes are
separated by regime shifts, which occurred about 1947, 1976-1977,
and 1989. These shifts are shown by the vertical dashed lines. The well-documented
1976-1977 regime shift has been associated with changing abundance
trends of other marine organisms such as plankton. It also has been related
to an abrupt change in the southernmost extent of sea ice in the Bering
Sea, in turn affecting the distribution of Walleye pollock (Theragra chalcogramma)
and Arctic cod (Boreogadus saida) (Wyllie-Echeverria and Wooster, 1998).
The concepts of regimes, regime shifts, and natural trends in the abundance
of animals indicates that it is important to assess the potential impact
of global warming on decadal-scale processes as well as on species specific
responses.
|
The collapse of the Peruvian anchovy (Engraulis ringens) fishery from
the mid-1970s to the mid-1980s was widely accepted as an example of overfishing
and poor management. The increases in recent years, to catches slightly smaller
than the large catches prior to the collapse, provide an example of the important
impact of natural fluctuations and the difficulty of sorting out the impacts
of fishing and climate-ocean-induced changes. Caddy and Rodhouse (1998) reported
an increase in world cephalopod landings as world marine fish catches stabilized.
These increases were believed to be related to reduced predation from overfishing
of groundfish stocks, although warmer oceans were considered an important factor.
These examples emphasize the importance of considering the ecosystem impacts
of climate variations, as well as changes for individual species.
McGowan et al. (1998) show that there are large-scale biological responses
in the ocean to climate variations. Off California, the climate-ocean regime
shift in 1976-1977 (Ebbesmeyer et al., 1991) resulted in a reduced
rate of supply of nutrients to a shallower mixing layer, decreasing productivity
and zooplankton and causing reductions in kelp and sea birds. Although there
is no question that fishing has impacts on the dynamics of fish populations,
the recent evidence of climate-related impacts is beginning to confound past
interpretations of fishing effects. McGowan et al. (1998) point out that
the success of future fish stock assessments would depend, to a large extent,
on the ability to predict the impacts of climate change on the dynamics of marine
ecosystems. The assumption that marine ecosystems are stable is no longer acceptable,
which raises questions about the definition of sustained yield (O'Brien
et al., 2000).
Weather impacts and seasonal rhythms have long been recognized by the global
fishing industry, but decadal-scale regime changes have been acknowledged only
recently as a factor in fish and ocean ecosystem dynamics. The concept of distinct
states in climate-ocean environments, which after periods of persistence switch
abruptly to other states, have been called regimes and regime shifts, respectively.
More formally, regimes (Steele, 1996) can be defined as multi-year periods of
linked recruitment patterns in fish populations or as a stable mean in physical
data. A regime shift is a change in the mean of a data series. The existence
of decadal-scale regimes in the environment has been documented (e.g., Gargett,
1997; Gu and Philander, 1997; Mantua et al., 1997). States even longer
than the decadal-scale may exist (Ware, 1995; Marsh et al., 1999). Adkinson
et al. (1996) and Beamish et al. (1997) have documented a large-scale
response in fish populations to regimes and regime shifts for Pacific salmon.
Among the most important groups of marine fishes are herrings (Clupea sp.),
sardines and pilchards (Sardinops sp.), and anchovies (Engraulis sp.)
(see Table 6-1). These fish tend to be short-lived species
that mature at an early age. Large fluctuations in abundance have been associated
with changes in the climate-ocean environment, although it has not been possible
to discover the mechanisms that link climate-ocean changes to recruitment (Cole
and McGlade, 1998). One of the most convincing relationships of large-scale,
synchronous responses in major fisheries resulting from changes in climate-ocean
states exists for sardine (Sardinops sp.). The decadal variability in
the Japanese sardine catch was synchronous with decadal-scale variability in
the ocean and climate of the North Pacific; these phenomena also were synchronous
with the fluctuations of sardine catches off Chile and California (Kawasaki,
1991; Hiyama et al., 1995) and with trends in Pacific salmon catches
(Beamish et al., 1999) (see Box 6-1).
| Table 6-1: Largest marine fisheries in 1996 (FAO,
1998). |
 |
| Species |
Landings (t)
|
% of Total
|
|
| Peruvian anchovy (Engraulis ringens) |
8,864,000
|
11.3
|
| Walleye pollock (Theragra chalcogramma) |
4,533,000
|
5.8
|
| Chilean Jack mackerel (Trachurus murphyi) |
4,379,000
|
5.8
|
| Atlantic herring (Clupea harengus) |
2,331,000
|
3.0
|
| Chub mackerel (Scomber japonicus) |
2,168,000
|
2.8
|
| Capelin (Mallotus villosus) |
1,527,000
|
1.9
|
| South American pilchard (Sardinops sagax) |
1,494,000
|
1.9
|
| Skipjack tuna (Katsuwonus pelamis) |
1,480,000
|
1.9
|
| Atlantic cod (Gadus morhua) |
1,329,000
|
1.7
|
| Largehead hairtail (Trichiarus lepturus) |
1,275,000
|
1.6
|
|
Fluctuations in the abundance and distribution of herring (Clupea harengus)
and sardine (Sadinella pilchardus) in the North and Baltic Seas have
been linked to variations of the North Atlantic Oscillation and the resulting
strength and pattern of southwesterly winds (Alheit and Hagen, 1997).
Most fishing regime changes can be related directly to sea-temperature changes,
but changes in other physical attributes also can have an impact. For instance,
a decrease in wind stress off Tasmania that reduced large zooplankton production
affected the density of Jack mackerel (Trachurus declivis), which eliminated
the possibility of a commercially viable mackerel fishery (Harris et al.,
1992).
|
Box 6-2. Tuna Migration and Climate Variability
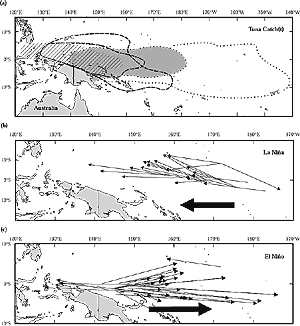
Skipjack tuna (Katsuwonus pelamis) dominate the world's catch of
tuna. The habitat supporting the densest concentrations of skipjack is
the western equatorial Pacific warm pool, with SST of 29°C and warmer.
Panel (a) clearly shows the association of skipjack tuna catch (shaded
and cross-hatched areas indicate January-June catch of 200,000+ t)
and mean SST [data from Lehodey et al. (1997)]. The figure also shows
that the location of the warm pool is linked to ENSO and that it changes
during El Niño and La Niña events. For instance, the catch
area in the first half of 1989 (La Niña period), which is shown
by cross-hatch, was centered around Palau and the Federated States of
Micronesia; in the first half of 1992 (El Niño period), the center
of abundance had shifted to the east, to the Marshall Islands and Kiribati
(shown by shading).
Panels (b) and (c) indicate the scale of tuna migration during a La Niña
period and an El Niño period, respectively. These figures were
compiled from records of a large-scale skipjack tagging program carried
out by the Secretariat of the Pacific Community (SPC). The illustrations
are from Lehodey et al. (1997, 1998).
The close association of skipjack tuna catch and ENSO is evidence that
climate variability profoundly affects the distribution pattern of tuna
and resulting fishing opportunities. Scientists do not know how projected
climate changes will affect the size and location of the warm pool in
the western and central Pacific, but if more El Niño-like conditions
occur an easterly shift in the center of tuna abundance may become more
persistent.
|
The aforementioned climate-related fluctuations in the Japanese sardine occurred
at the same time as shifts in the migratory patterns of the northern bluefin
tuna (Thunnus thynnus), with a higher proportion of bluefin remaining
in the western Pacific when sardine abundance was high (Polovina, 1996). The
migratory pattern of albacore tuna (T. alalunga) also was altered by decadal-scale
climate changes (Kimura et al., 1997).
Nearly 70% of the world's annual tuna harvest comes from the Pacific Ocean.
In 1996, Skipjack tuna (Katsuwonus pelamis) was the eighth-largest marine
fishery in the world (see Table 6-1). Catches of skipjack
are highest in the western equatorial Pacific warm pool; Lehodey et al.
(1997) have shown that major spatial shifts in the skipjack population can be
linked to large zonal displacements of the warm pool (see Box
6-2).
Welch et al. (1998) propose that continued warming of the North Pacific
Ocean would compress the distributions of Sockeye salmon (Oncorhynchus nerka),
essentially squeezing them out of the North Pacific and into the Bering Sea.
Some modeling of future impacts of greenhouse gas increases, however, has shown
an intensification of the Aleutian Low, which has been associated with mid-ocean
cooling (Deser et al., 1996) and increased Pacific salmon production.
The warmer surface waters could reduce growth if bioenergetic costs are higher
and less food is available as a consequence of ocean habitat changes. The potential
impact of climate change on Pacific salmon is expected to occur in freshwater
and ocean situations (Hinch et al., 1995). This fact is important because
production of more juveniles in hatcheries would not mitigate changes in the
ocean carrying capacity for Pacific salmon. The most effective strategy to manage
the impacts of climate change on Pacific salmon may be to ensure that wild salmon
are preserved and protected, rather than to produce more salmon through artificial
enhancement. It is possible that the variety of life history types and genetic
traits of the wild stocks are inherent biological solutions to changing freshwater
and marine habitats (Bisbal and McConnaha, 1998).
The potential deepening of the Aleutian Low and increase in the amplitude of
the Pacific Decadal Oscillation would result in major changes in marine ecosystems
(Mantua et al., 1997). As ecosystems change, there may be impacts on
the distribution and survival of fishes. Any changes in natural mortality would
be associated with increased predation and other factors such as disease. Improved
growth in the early life stages would improve survival, whereas decreased growth
could facilitate increased mortality.
Sea temperature is an important regulator of fish behaviors. Wood and McDonald
(1997) provide examples of how climate change could induce temperature responses
in fish, but there are several areas where less certainty exists. The effect
that global climate change will have on trends in the Aleutian Low Pressure
system in the Pacific Ocean is an example. Although there are clear linkages
between the intensity and position of the low and production trends of many
of the commercially important fish species (Kawasaki et al., 1991; Polovina
et al., 1995; Gargett, 1997; Mantua et al., 1997; Francis et
al., 1998), a reduction in equator-to-pole temperature gradients would probably
weaken winds and consequently reduce open-ocean upwelling. Important changes
in species distributions in surface waters could result.
There is now a cautious acceptance that climate change will have major positive
and negative impacts on the abundance and distribution of marine fish. Thus,
the impacts of fishing and climate change will affect the dynamics of fish and
shellfish such as abalone in Mexico (Shepherd et al., 1998). Fishing
impacts may be particularly harmful if natural declines in productivity occur
without corresponding reductions in exploitation rates. Changes in fish distributions
and the development of aquaculture may reduce the value of some species, however—as
it has for wild Pacific salmon—and these changes may reduce fishing pressures
in some areas.
Key to understanding the direction of change for world fisheries is the ability
to incorporate decadal-scale variability into general circulation models (GCMs).
Although progress has occurred, it still is not possible to assess regional
responses to shifts in climate trends, and it is unknown if a general warming
will increase or decrease the frequency and intensity of decadal-scale changes
in regions where national fisheries occur. Recent studies have not produced
evidence to change the conclusion from the SAR (Everett et al., 1996)
that future saltwater fisheries production is likely to be about the same as
at present, though changes in distribution could affect who catches a particular
stock. However, if aquaculture becomes the major source of fish flesh and management
of fisheries becomes more precautionary, the exploitation rate of wild marine
fish may decrease in some areas.